Deoxyribonucleic acid - DNA - serves as a carrier of hereditary information transmitted by living organisms to next generations, and as a matrix for the construction of proteins and various regulatory factors that are needed by the body in the processes of growth and vital activity. In this article, we will focus on what are the most common forms of DNA structure. We will also pay attention to how these forms are built and in what form the DNA resides inside a living cell.
DNA molecule organization levels
There are four levels that determine the structure and morphology of this giant molecule:
- The primary level, or structure, is the order of the nucleotides in the chain.
- The secondary structure is the famous “double helix”. It was this phrase that was established, although in fact, such a structure resembles a screw.
- The tertiary structure is formed due to the fact that between the individual sections of the double-stranded twisted DNA strand weak hydrogen bonds arise, giving the molecule a complex spatial conformation.
- The quaternary structure is already a complex complex of DNA with some proteins and RNA. In this configuration, the DNA is packed into chromosomes in the cell nucleus.
Primary structure: DNA components
The blocks from which the deoxyribonucleic acid macromolecule is built are nucleotides, which are compounds, each of which includes:
- nitrogenous base - adenine, guanine, thymine or cytosine. Adenine and guanine belong to the group of purine bases, cytosine and thymine - pyrimidine;
- five-carbon deoxyribose monosaccharide;
- phosphoric acid residue.
In the formation of a polynucleotide chain, an important role is played by the order of the groups formed by carbon atoms in the ring sugar molecule. The phosphate residue in the nucleotide is connected to the 5′-group (read “five prime”) of deoxyribose, that is, to the fifth carbon atom. The chain extension occurs by adding to the free 3'-group of deoxyribose phosphate residue of the next nucleotide.
Thus, the primary structure of DNA in the form of a polynucleotide chain - has 3'- and 5'-ends. This property of a DNA molecule is called polarity: chain synthesis can only go in one direction.
Secondary structure formation
The next step in the structural organization of DNA is based on the principle of complementarity of nitrogenous bases - their ability to pair with each other through hydrogen bonds. Complementarity - mutual correspondence - arises because adenine and thymine form a double bond, and guanine and cytosine form a triple bond. Therefore, when forming a double chain, these bases stand opposite each other, forming the corresponding pairs.
Polynucleotide sequences are antiparallel in the secondary structure. So, if one of the chains has the form 3 '- AHGTSATAA - 5', then the opposite will look like this: 3 '- TTATGSTST - 5'.
When a DNA molecule is formed, twisting of the double polynucleotide chain takes place, and what forms the DNA can take at this structural stage depends on the concentration of salts, on water saturation, on the structure of the macromolecule itself. Several such forms are known, denoted by the Latin letters A, B, C, D, E, Z.
Configurations C, D, and E were not found in wildlife and were observed only in laboratory conditions. We will consider the main forms of DNA: the so-called canonical A and B, as well as the configuration Z.
A-DNA is a dry molecule
An A-shape is a right-handed screw with 11 complementary base pairs in each turn. Its diameter is 2.3 nm, and the length of one coil of the spiral is 2.5 nm. The planes formed by paired bases have an inclination of 20 ° with respect to the axis of the molecule. Neighboring nucleotides are compactly arranged in chains - between them only 0.23 nm.
This form of DNA occurs with low hydration and with an increased ionic concentration of sodium and potassium. It is characteristic of processes in which DNA forms a complex with RNA, since the latter is not able to take other forms. In addition, the A-form is very resistant to ultraviolet radiation. In this configuration, deoxyribonucleic acid is found in fungal spores.
Wet B-DNA
With a low salt content and a high degree of hydration, that is, under normal physiological conditions, DNA takes its main form B. Natural molecules exist, as a rule, in the B-form. It is she who underlies the classic Watson-Crick model and is most often depicted in the illustrations.
This form (it is also right-handed) is characterized by a smaller compactness of nucleotide placement (0.33 nm) and a large screw pitch (3.3 nm). One turn contains 10.5 base pairs, the rotation of each of them relative to the previous one is about 36 °. The plane of the pairs is almost perpendicular to the axis of the "double helix". The diameter of such a double chain is smaller than that of the A-form - it reaches only 2 nm.
Noncanonical Z-DNA
Unlike canonical DNA, the Z-type molecule is a left-handed screw. It is the thinnest of all, has a diameter of only 1.8 nm. Its coils of 4.5 nm in length are as if elongated; this form of DNA contains 12 paired bases per coil. The distance between adjacent nucleotides is also quite large - 0.38 nm. So the Z-shape is characterized by the least degree of torsion.
It is formed from a B-type configuration in those areas where purine and pyrimidine bases alternate in the nucleotide sequence when the content of ions in the solution changes. The formation of Z-DNA is associated with biological activity and is a very short-term process. This form is unstable, which creates difficulties in the study of its functions. So far, they are not exactly clear.
DNA replication and its structure
Both the primary and secondary structures of DNA arise in the course of a phenomenon called replication — the formation of two “double helices” identical to it from the maternal macromolecule. During replication, the original molecule unravels, and complementary bases build up on the released single chains. Since the halves of DNA are antiparallel, this process proceeds in them in different directions: in relation to the mother chains from the 3'-end to the 5'-end, that is, new chains grow in the direction 5 '→ 3'. The leading chain is synthesized continuously towards the replication fork; on the lagging chain, synthesis takes place from the plug in separate sections (Okazaki fragments), which then cross-link a special enzyme, DNA ligase.
As synthesis continues, the already formed ends of the daughter molecules undergo helical twisting. Then, even before the end of replication, newborn molecules begin to form a tertiary structure in a process called supercoiling.
Supercoiled molecule
The supercoiled form of DNA occurs when a double-stranded molecule performs an extra twist. It can be directed clockwise (positively) or against (in this case, they speak of negative supercoiling). The DNA of most organisms is supercoiled negatively, that is, against the main turns of the “double helix”.
As a result of the formation of additional loops - supercoils - DNA acquires a complex spatial configuration. In eukaryotic cells, this process occurs with the formation of complexes in which DNA is negatively wound on histone protein complexes and takes the form of a strand with bead nucleosomes. Free sections of the thread are called linkers. Non- histone proteins as well as inorganic compounds take part in maintaining the supercoiled form of the DNA molecule. This is how chromatin is formed - the substance of chromosomes.
Chromatin filaments with nucleosome beads are capable of further complicating the morphology in a process called chromatin condensation.
DNA final compaction
In the nucleus, the form of the deoxyribonucleic acid macromolecule becomes extremely complex, compacting in several stages.
- Firstly, the thread is folded into a special structure such as a solenoid — a chromatin fibril 30 nm thick. At this level, DNA, folding, reduces its length by 6-10 times.
- Further, the fibril with the help of specific scaffold proteins forms zigzag loops, which reduces the linear size of DNA by 20-30 times.
- At the next level, tightly packed looped domains are formed, most often having the shape conventionally called the “tube brush”. They attach to the intranuclear protein matrix. The thickness of such structures is already 700 nm, while DNA is shortened by about 200 times.
- The last level of morphological organization is chromosomal. Loop domains are compacted so that a total shortening of 10,000 times is achieved. If the length of the stretched molecule is about 5 cm, then after packing in the chromosomes it decreases to 5 microns.
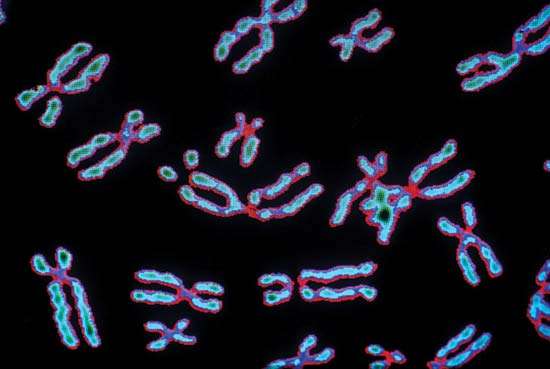
The highest level of complication of the form of DNA is reached in the metaphase state of mitosis. It was then that it acquires a characteristic appearance - two chromatids connected by a centromere constriction, which ensures the separation of chromatids in the fission process. Interphase DNA is organized to the domain level and is distributed in the cell nucleus without any special order. Thus, we see that the morphology of DNA is closely related to the various phases of its existence and reflects the functioning of this molecule, which is most important for life.