Nuclear pores are one of the most important intracellular components, since they are involved in molecular transport. Despite advances in biological research, not all issues concerning these structures are fully understood. Some scientists believe that by the importance of functions and structural complexity, the complex of nuclear pores can be attributed to cell organelles.
Nuclear shell
A characteristic feature of eukaryotic cells is the presence of a nucleus, which is surrounded by a membrane that separates it from the cytoplasm. The membrane consists of two layers - internal and external, interconnected using a large number of pores.
The value of the nuclear envelope is very high - it allows you to delimit the processes of protein synthesis and nucleic acids necessary for regulating the functional activity of genes. The membrane controls the process of transporting substances inward, into the cytoplasm, and in the opposite direction. It is also a skeletal structure supporting the shape of the nucleus.
Between the outer and inner membranes is the perinuclear space, the width of which is 20-40 nm. Externally, the nuclear shell looks like a two-layer bag. The presence of pores in its structure is a significant difference between this structure and those of mitochondria and plastids.
The structure of nuclear pores
The channels are perforations with a diameter of about 100 nm, passing through the entire nuclear shell. In cross section, they are characterized by the shape of a polygon having eighth-order symmetry. The substance-permeable channel is in the center. It is filled with complexly organized globular (in the form of a ball) and fibrillar (in the form of a twisted thread) structures that form the central “plug” granule (or conveyor). In the figure below, you can visually examine what a nuclear pore is.
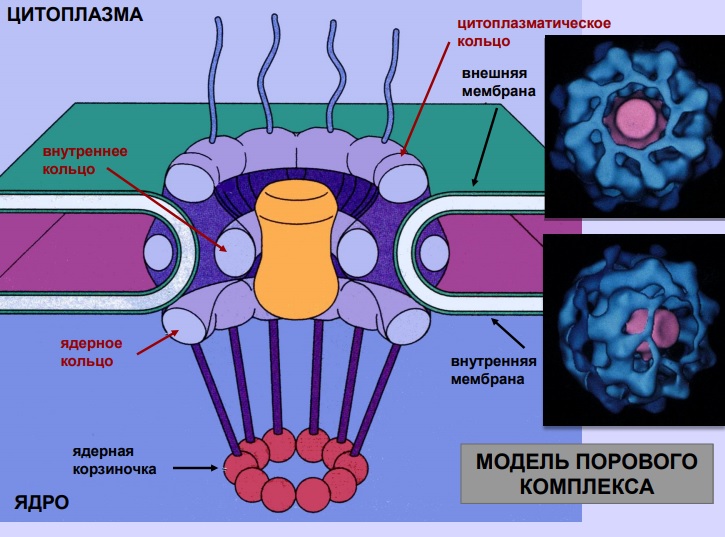
Microscopic examination of these structures shows that they have a ring structure. Fibrillar outgrowths extend both outward, into the cytoplasm, and inward, towards the nucleus (filaments). The latter form a kind of basket (called “basket” in foreign literature). In the passive pore, the fibrils of the basket close the channel, and in the active pore, an additional formation with a diameter of about 50 nm is formed. The ring from the side of the cytoplasm consists of 8 granules interconnected like beads on a thread.
The totality of these perforations in the shell of the nucleus is called the complex of nuclear pores. Thus, biologists emphasize the interconnectedness of individual holes working as a single cohesive mechanism.
The outer ring is connected to the central conveyor. Lower eukaryotes (lichens and others) do not have cytoplasmic and nucleoplasmic rings.
Structure Features
The structure and functions of nuclear pores have the following features:
- Channels are numerous copies of the order of 30-50 nucleoporins (and a total of about 1000 proteins).
- The mass of complexes ranges from 44 MDa in lower eukaryotes to 125 MDa in vertebrates.
- In all organisms (humans, birds, reptiles and other animals) in all cells, these structures are arranged in a similar way, that is, pore complexes are strictly conservative.
- The components of nuclear complexes have a subunit structure, due to which they have high plasticity.
- The diameter of the central channel varies between 10-26 nm, and the height of the pore complex is about 75 nm.
The nuclear pore regions remote from the center are asymmetric. Scientists attribute this to various mechanisms for regulating transport function in the initial stages of cell development. It is also assumed that all pores are universal structures and provide the movement of molecules both in the cytoplasm and in the opposite direction. Nuclear pore complexes are also present in other components of the cell that have membranes, but in more rare cases (reticulum, terminal membranes of the cytoplasm).
Number of pores
The main factor on what the number of nuclear pores depends on is the metabolic activity in the cell (the higher it is, the greater the number of tubules). Their concentration in the thickness of the membrane can vary several times at different periods of the functional state of the cells. The first increase in the number of pores occurs after division - mitosis (during the reconstruction of nuclei), and then during the period of DNA growth.
In different species of animals, their number is different. It also depends on where the sample was taken. So, in the culture of human tissues , there are about 11 pcs / μm 2 , and in the unripened egg cell of a xenopus frog - 51 pcs / μm 2 . On average, their density varies between 13-30 pieces / μm 2 .
The distribution of nuclear pores over the surface of the shell is almost uniform, but in the places where the substance of chromosomes approaches the membrane, their concentration decreases sharply. Lower eukaryotes under the nuclear membrane do not have a fibrillar network of a rigid structure; therefore, pores can move along the nuclear envelope, and their density at different sites varies significantly.
Functions
The main function of the nuclear pore complex is the passive (diffusion) and active (requiring energy costs) transfer of molecules through the membrane, that is, the metabolism between the cell nucleus and the cytoplasm. This process is vital and is regulated by three systems that are in constant interaction with each other:
- a complex of biologically active substances-regulators in the nucleus and cytoplasm - importin α and β, Ran-protein, guanosine triphosphate (purine nucleotide) and other inhibitors and activators;
- nucleoporins;
- structural components of the pore nuclear complex, which are able to change their shape and ensure the transfer of substances in the right direction.
Proteins necessary for the functioning of the nucleus come from the cytoplasm through the nuclear pores, and various forms of RNA are removed in the opposite direction. The pore complex not only carries out purely mechanical transportation, but also serves as a sorter, “recognizing” certain molecules.
Passive transmission occurs for those substances whose molecular weight is low (not more than 5 ∙ 10 3 Yes). Such substances as ions, sugars, hormones, nucleotides, adenosine triphosphoric acid, participating in the energy exchange, freely enter the nucleus. The maximum size of proteins that can penetrate through the pores into the nucleus is 3.5 nm.
During the synthesis of a daughter DNA molecule, the transportation of substances reaches a peak of activity - 100-500 molecules through 1 nuclear pore in 1 min.
Protein Pores
Elements of the channels are protein in nature. The proteins of this complex are called nucleoporins. They are collected in approximately 12 subcomplexes. They are conditionally divided into three groups:
- compounds with specific repeating sequences recognized by biochemical factors;
- not having sequences;
- integral proteins that are located in the region of the membrane that forms the pore, or in the pore itself in the space between the layers of the nuclear membrane.
Studies have shown that nucleoporins can form quite complex complexes, including up to 7 proteins, and also take a direct part in the transport of substances. Some of them can directly bind to molecules moving through the nuclear pore.
Export of substances into the cytoplasm
One and the same time can take part both in the withdrawal and in the import of substances. The reverse transfer of RNA from the cytoplasm to the nucleus does not occur. Nuclear complexes recognize export signals (NES) that carry ribonucleoproteins.
The NES sequence of signaling substances is a complex complex of amino acids and proteins, which, after being excreted from the nucleus into the cytoplasm, dissociate (decompose into individual components). Therefore, similar particles introduced into the cytoplasm by artificial means do not penetrate back into the nucleus.
Mitosis process
During cell division (mitosis), the nuclear pore complex is “disassembled”. Thus, complexes with a molecular mass of 120 mDa decompose into subcomplexes of 1 mDa each. After the end of the division, they reassemble. At the same time, nuclear pores are not moved separately, but by arrays. This is one of the evidence that the nuclear pore complex is a coherent system.
The destroyed membrane turns into a vesicle cluster that surrounds the core region during the interphase period. In metaphase, when chromosomes are held in the equatorial plane, these elements are pushed to the peripheral zones of the cell. At the end of anaphase, this cluster begins to contact the chromosomes and the growth of the beginnings of the nuclear membrane starts.
Bubbles turn into vacuoles, which gradually envelop the chromosomes. Then they merge and block the new interphase nucleus from the cytoplasm. Pores appear already at an early stage, when the closure of the shells has not yet occurred.